Leksjoner som ikke er gått hjem i den evangeliske debatten over Adam og Eva.
Oversatt herfra. Du finner de tidligere tekstene, nummerert under samme tittel, her.
Bilde 1. Paret i fokus: Adam og Eva
 I den forrige delen av min anmeldelse av William Lane Craigs 'In Quest of the Historical Adam', så vi at mange evangeliske intellektuelle hadde akseptert argumenter om at Adam og Eva ikke kunne ha eksistert. Disse argumentene, spesielt påstanden om at menneskets genetiske mangfold er for stort, til å ha blitt redusert til et enkelt par, ble kraftig fremmet av teistiske evolusjonister aka evolusjonære kreasjonister (TE/ECs) tilknyttet BioLogos. Fremtredende blant disse kritikerne var Dennis Venema, en biolog ved Trinity Western University, som sammenlignet dagens tro på Adam og Eva med å følge den lenge tilbakeviste geosentriske modellen av solsystemet. Men argumentene viste seg å være feil, som til og med BioLogos og Venema nå innrømmer.
I den forrige delen av min anmeldelse av William Lane Craigs 'In Quest of the Historical Adam', så vi at mange evangeliske intellektuelle hadde akseptert argumenter om at Adam og Eva ikke kunne ha eksistert. Disse argumentene, spesielt påstanden om at menneskets genetiske mangfold er for stort, til å ha blitt redusert til et enkelt par, ble kraftig fremmet av teistiske evolusjonister aka evolusjonære kreasjonister (TE/ECs) tilknyttet BioLogos. Fremtredende blant disse kritikerne var Dennis Venema, en biolog ved Trinity Western University, som sammenlignet dagens tro på Adam og Eva med å følge den lenge tilbakeviste geosentriske modellen av solsystemet. Men argumentene viste seg å være feil, som til og med BioLogos og Venema nå innrømmer.
Til sin kredit er William Lane Craig blant de evangelikale som har vært villige til å stille spørsmål ved argumenter mot Adam og Eva. I sin bok siterer han arbeidet til Ann Gauger, Ola Hössjer og Joshua Swamidass som utførte analyser, som viste at menneskeheten kunne ha oppstått fra et enkelt par for minst 500 000 år siden. Gauger og Hössjer bemerket at Adam og Eva kunne ha levd enda nyere, hvis ytterligere evolusjonære antakelser blir stilt spørsmål ved.
Da jeg leste retorikken brukt av evangelikale eliter som tok til orde for å forlate en historisk Adam og Eva, ble jeg slått av hvor mye av den, som virket drevet av frykt - frykt for å se tåpelig ut foran verden, fordi du utfordret evolusjonen og ble vist å ta feil. Som jeg diskuterte, er lærdommen fra denne historien at det bør være akseptabelt å utfordre evolusjonære argumenter. Vi trenger ikke leve i frykt for at å gjøre det er 'antivitenskap' eller vil "bringe vanry over den kristne tro" eller "skam over Jesu Kristi navn" - som noen evangelkale eliter har hevdet.
I denne siste delen av anmeldelsen min vil vi se at dessverre har noen fremtredende kristne ikke fullt ut satt pris på denne leksjonen. De virker innstilt på å holde seg til konsensus, gjenta tidligere feil og omfavne evolusjonære ideer - til tross for mangel på overbevisende bevis. For å være spesifikk:
1. BioLogos, Joshua Swamidass og (noen ganger) William Lane Craig forfekter nå en spesiell modell av menneskelig opprinnelse der vi alle stammer fra en populasjon av hominider som utviklet seg fra en felles stamfar vi deler med aper, via standard evolusjonsmekanismer - akkurat som evolusjonsteorien foreslår. Under denne 'genealogiske Adam og Eva'-modellen var Adam og Eva historiske mennesker som kan ha blitt spesielt skapt, men deres avkom blandet seg med de fullt utviklede hominidene, noe som til slutt førte til oss. Denne modellen er fundamentalt evolusjonær, men bevisene motsier den, eller krever det ikke.
Bilde 2. Han mener de eksisterte, men hvordan? (W.L. Craig)
 2. Craig ser i boken sin, til tider ut til å misforstå modellen ovenfor,
2. Craig ser i boken sin, til tider ut til å misforstå modellen ovenfor,
Fordi den antyder "dyriske forhold til ikke-menneskelige homininer." Han foretrekker å si at "Ingen slik appell til kryssing er nødvendig, hvis vi ser for oss at Adam og Eva kommer fra en homininpopulasjon som delte felles aner med sjimpanser og andre menneskeaper." (s. 378) Denne modellen utfordres også av bevisene.
3. Enten Craig støtter en standard evolusjonsmodell der Adam og Eva er direkte utviklet fra en felles stamfar som deles med aper (modell 2 ovenfor), eller den nye modellen der Adam og Evas avkom blandet seg og blandet med hominider som utviklet seg fra en felles stamfar delt med aper (modell 1 ovenfor), blir Craig drevet av ideen om at mennesker deler en felles stamfar med aper. I begge tilfeller siterer han 'ødelagte' pseudogener som bevis for menneske-apers felles aner - og stoler på Dennis Venema som en primær kilde. Den beste tilgjengelige vitenskapen krever heller ikke at denne modellen er sann.
I modellene ovenfor ser det ut til at noen kristne ikke fullt ut har satt pris på lærdommen fra evangeliske debatter om Adam og Eva (Del 4): Vi trenger ikke frykte at det å utfordre en evolusjonsmodell nødvendigvis strider mot god vitenskap. Faktisk, noen ganger kan det å beholde en evolusjonsmodell forårsake selve frykten den var ment å avverge - få oss til å se dårlig ut foran verden fordi vi utfordret vitenskapen og tok feil. I stedet for frykt, la oss drives av et ønske om å finne sannheten. Så la oss hugge inn.
Bilde 3. Ikke bare protein-kodende gener er virksomme
 Ikke å lære leksjonen om den genealogiske Adam og Eva-modellen
Ikke å lære leksjonen om den genealogiske Adam og Eva-modellen
En artikkel fra BioLogos fra juli 2021 erkjenner nå muligheten for en versjon av en historisk Adam og Eva -lenke . De sier:
Alternativene inkluderer .. postulerer at Adam og Eva var et spesielt, historisk par som faktisk var forfedre til alle som lever i dag, men som var en del av en større befolkning som deres etterkommere kunne parre seg med. (1)
Under dette synet var Adam og Eva historiske personer hvis avkom blandet seg med en populasjon av hominider som utviklet seg fra apelignende forfedre. Merk imidlertid at BioLogos skiller dette synet fra det det kaller det "vanlige tradisjonelle" synet der "Adam og Eva ble skapt de novo" som våre "eneste forfedre": De var de to første menneskene, og de alene ga opphav til alle andre mennesker.-"(2) Det ser ut til at BioLogos fortsatt misforstår dette tradisjonelle synet, og foretrekker i stedet at Adam og Eva eksisterte, men at deres avkom blandet seg med en stor populasjon av mennesker som utviklet seg naturlig fra apelignende skapninger. Derfor foreslår BioLogos i dag tilsynelatende at vi stammer fra både Adam og Eva så vel som utviklede hominider - og vi deler en felles stamfar med levende aper. Denne "Genealogical Adam and Eva" (GAE) modellen er definitivt ikke det 'vanlige tradisjonelle' synet.
GAE-modellen ble utdypet av Joshua Swamidass i hans bok fra 2019 The Genealogical Adam and Eve.(3) Som vi så i del 4 av denne anmeldelsen, tar William Lane Craig til tider avstand fra dette forslaget fordi det innebærer 'dyriske' forhold mellom etterkommerne av Adam og Eva og ikke-menneskelige homininer. Imidlertid ser det ut til at han, i siste instans, tillater GAE-modellen.
Bilde 4. Bakgrunnen dreier seg om hvorvidt et darwinistisk scenario er realistisk
 Som kristen tror jeg det er alvorlige teologiske og skriftmessige problemer med GAE-modellen, men dette er ikke stedet for slike argumenter. I alle fall er mine hovedinnvendinger vitenskapelige. Vitenskapelig tar GAE-modellen i bruk et standard evolusjonært syn på menneskelig opprinnelse og sier at hvis Adam og Eva ble spesielt skapt, ble deres avkom fullstendig blandet med en fullt utviklet populasjon av hominider. Vi er etterkommere av denne store befolkningen. Fordi GAE tar i bruk en standard evolusjonær beretning om menneskelig opprinnelse, arves eventuelle vitenskapelige problemer med en slik beretning av GAE-modellen. Er de vitenskapelige bevisene så overbevisende at vi må akseptere dette synet?
Som kristen tror jeg det er alvorlige teologiske og skriftmessige problemer med GAE-modellen, men dette er ikke stedet for slike argumenter. I alle fall er mine hovedinnvendinger vitenskapelige. Vitenskapelig tar GAE-modellen i bruk et standard evolusjonært syn på menneskelig opprinnelse og sier at hvis Adam og Eva ble spesielt skapt, ble deres avkom fullstendig blandet med en fullt utviklet populasjon av hominider. Vi er etterkommere av denne store befolkningen. Fordi GAE tar i bruk en standard evolusjonær beretning om menneskelig opprinnelse, arves eventuelle vitenskapelige problemer med en slik beretning av GAE-modellen. Er de vitenskapelige bevisene så overbevisende at vi må akseptere dette synet?
Som vi så i del 3 -lenke av denne anmeldelsen, er de fossile bevisene for menneskelig evolusjon fra apelignende skapninger svake, og neo-darwinistiske mekanismer står overfor en overveldende matematisk hindring for å forklare opprinnelsen til komplekse menneskelige egenskaper som våre kognitive evner. Av disse grunnene er mange aspekter av GAE-modellen vitenskapelig problematiske. Hvis mennesker ikke utviklet seg fra apelignende skapninger via standard evolusjonsmekanismer, ser jeg ingen tvingende grunn til å vedta GAE-hypotesen.
Videre, i del 4 av denne gjennomgangen -lenke, så vi at Gauger og Hössjers forskning - så vel som Swamidass egen modellering - viser at hvis Adam og Eva levde langt nok tilbake i fortiden, så er moderne menneskelig genetisk mangfold forenlig med et første par som var våre eneste forfedre. Dette eliminerer ethvert behov for å påkalle tusenvis av evolusjonære forfedre, som er en nøkkelfunksjon i GAE-modellen.
Så hvis det vitenskapelige beviset for menneskelig evolusjon er svakt og GAE-modellen ikke engang nødvendigvis kreves av bevisene, hvorfor støttes den av BioLogos, Swamidass og (det ser ut til, til slutt) William Lane Craig? Dette er komplekse spørsmål, og det er sannsynligvis mange årsaker. Men hvis jeg måtte spekulere, er noen av dem sannsynligvis de samme grunnene som førte til at folk feilaktig avviste en historisk Adam og Eva i utgangspunktet.
GAE-modellen lar en beholde troen på en versjon (om enn en utradisjonell) av en historisk Adam og Eva, mens man fortsatt tar i bruk en fullstendig evolusjonær modell av menneskelig opprinnelse. Dette appellerer til dem som (som vi så i del 4 av denne anmeldelsen) feilaktig tror at å utfordre evolusjon "bringer vanry over den kristne tro-" og "unødvendig skam over Jesu Kristi navn-". Paul Nelson har overbevisende hevdet at den bærende filosofien bak GAE-modellen er en forpliktelse til metodologisk naturalisme (4), ideen om at når man studerer vitenskap, har man kun lov til å påberope seg naturalistiske krefter og mekanismer. Kanskje ikke alle GAE-tilhengere føler sterkt for metodisk naturalisme, men de føler absolutt sterkt for å beholde en evolusjonær modell.
Men hvis vi bryr oss mer om vitenskap og sannhet enn om evolusjon, metodologisk naturalisme eller verdslig aksept, så er det kanskje GAE-modellen som er 'unødvendig'. Jeg er sikker på at mange GAE-tilhengere vil være uenige og si at det kreves av vitenskapen - men er vitenskapen virkelig så klar på at vi må dele en felles stamfar med aper? Vi kommer til bevisene snart. {I mellomtiden kan en undres overr manglende bevis her -oversetters kommentar.}
Uansett årsakene til å ta i bruk den Genealogiske Adam og Eva-modellen, som BioLogos innrømmer, er det ikke den 'vanlige tradisjonelle' Adam og Eva. Under GAE-scenarioet er ikke Adam og Eva menneskehetens eneste forfedre, og vi stammer ikke bare fra dem, men også fra en stor populasjon av mennesker som fullt ut utviklet seg fra apelignende forfedre. Dermed vil kristne som holder seg til en tradisjonell Adam og Eva ha problemer med GAE-modellen.
Bilde 5. Enten felles design eller felles avstamning
Ikke-lært leksjon om pseudogener
 Selv om Craig forteller historien om bortfallet av BioLogos sine argumenter mot Adam og Eva, bruker han fortsatt argumentene deres for å støtte felles aner. Med henvisning til Dennis Venema, nevner Craig pseudogener som bevis for felles menneske-ape aner:
Selv om Craig forteller historien om bortfallet av BioLogos sine argumenter mot Adam og Eva, bruker han fortsatt argumentene deres for å støtte felles aner. Med henvisning til Dennis Venema, nevner Craig pseudogener som bevis for felles menneske-ape aner:
Man kan, med Swamidass og Hössjer og Gauger, i stedet postulere en de novo-skapelse av Adam og Eva. Men så står man overfor et vanskelig dilemma. Man må forklare vår genetiske likhet med sjimpanser enten på grunnlag av gjentatt guddommelig bruk av en lignende designplan eller på grunnlag av betydelig avling med ikke-mennesker. Den første har problemer med å forklare ødelagte pseudogener som vi deler med sjimpanser ... (s. 376, uthevelse tilføyd).
Som nevnt, avviser Craig til tider GAE-modellen fordi den innebærer bestialitet mellom etterkommere av Adam og Eva og ikke-menneskelige hominider. Hvis han ikke aksepterer GAE-modellen, og vil at Adam og Eva skal være våre eneste genetiske stamfar, har han tilsynelatende to valg: godta en versjon av Adam og Eva der de på mirakuløst vis er skapt og vi ikke deler en felles stamfar med aper, eller se på Adam og Eva som fullt utviklede homininer som delte en felles stamfar med aper. Craig foretrekker tilsynelatende det siste alternativet - felles aner:
Noen har appellert til avl med andre utviklede homininarter for å forklare hvordan Adam og Eva kunne ha vært de novo-skapelser fra livløst materiale, og likevel har deres etterkommere en så slående genetisk likhet med sjimpanser, inkludert ødelagte pseudogener som har opphørt med sin opprinnelige funksjon (S. Joshua Swamidass, The Genealogical Adam and Eva: The Surprising Science of Universal Ancestry [Downers Grove, IL: IVP Academic, 2019]). Jfr. Dennis Venemas kritikk av Hössjer og Gaugers de novo kreasjonisme med den begrunnelse at det menneskelige genomet er fylt med bevis på at vi deler felles forfedre med andre arter, som sjimpanser og gorillaer (Dennis Venema, "Adam-Once More, with Feeling-", Jesus Creed (blogg), 4. november 2019, -lenke). Ingen slik appell til kryssing er nødvendig hvis vi ser for oss at Adam og Eva kommer fra en homininpopulasjon som delte felles aner med sjimpanser og andre menneskeaper. Faktisk, på det synet som er foreslått her, kan Adam og Eva være våre eneste genetiske forfedre, hvis etterkommere aldri falt i dyriske forhold med ikke-menneskelige homininer eller i det minste ikke produserte noen etterkommere fra slike forbindelser. (s. 378)
Som vi så i del 4 av denne anmeldelsen, er GAE-modellen designet for å beholde menneske-ape felles aner og gi rom for en mirakuløst skapt historisk Adam og Eva, men den mister Adam og Eva som våre "eneste genetiske stamfedre." Den direkte felles forfedremodellen, som Craig ser ut til å foretrekke ovenfor, ser på Adam og Eva som etterkommere fra en felles stamfar som deles med aper, men tillater ikke at de blir "mirakuløst skapt de novo." Begge modellene involverer menneske-ape felles aner, og selv om Craig ikke er klar over hvilken modell han foretrekker, er det klart at han foretrekker en versjon av en menneske-sjimpanse felles anermodell fremfor en virkelig tradisjonell modell av Adam og Eva der de er våre eneste genetiske forfedre, ble mirakuløst skapt de novo, og ikke deler felles aner med aper.
Hvorfor er Craig villig til å gi opp kjerneaspekter ved en tradisjonell Adam og Eva til fordel for en modell som krever felles aner fra menneske-aper? Svaret kan oppsummeres i to ord, som han bruker om begge sitatene ovenfor: "ødelagte pseudogener-".
Logikken går som følger: Pseudogener er ødelagte gener som en gang hadde funksjon, men som ble inaktivert av en eller annen mutasjon. Gud ville aldri lagt ødelagt DNA i to arter på samme sted, lyder argumentet. Derfor, hvis mennesker og aper deler ikke-funksjonelle pseudogener i samme plassering av våre respektive genomer, må vi ha ervervet disse pseudogenene gjennom en naturlig ikke-designet mekanisme - nemlig arv fra en felles stamfar.
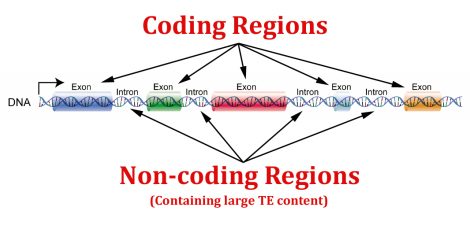
Intelligent design er kompatibelt med felles opphav, så når vi diskuterer dette emnet, tester vi ikke nødvendigvis ID på makroskala. Men i liten skala synes logikken riktig, at ødelagt DNA som deles av to arter er bedre forklart av materielle evolusjonære årsaker enn av intelligent årsakssammenheng. Men hvis pseudogener ikke er ødelagt, ikke-funksjonelt søppel-DNA, øker dette muligheten for at de er viktige funksjonelle deler av genomet vårt. I så fall er grunnen til at vi deler "pseudogent" DNA med aper ikke felles aner, men av funksjonelt viktige årsaker som gjenspeiler felles design.
Så er pseudogener virkelig "ødelagt" og ikke-funksjonelt søppel-DNA? Vel, hva sier litteraturen?
Bilde 6. Stadig flere kjente funksjoner til 'junk-DNA'
En raskt voksende trend i teknisk litteratur: Pseudogen funksjon
Faktisk er antallet artikler som rapporterer omfattende funksjon for pseudogener imponerende. Mange fremtredende fagfellevurderte vitenskapelige artikler har sagt ting som (min parafrase): "Vi pleide å tro at pseudogener var søppel-DNA, men dette er bare fordi vi egentlig ikke hadde teknologien til å forstå dem. Vi forstår fortsatt knapt pseudogener. Men nå som vi utvikler metoder for å studere dem, oppdager vi at funksjonalitet er ganske vanlig.-" Dette betyr at det kan være klokere å ta i bruk en "vent og se"-tilnærming til pseudogener, og ikke for tidlig konkludere med at de bare er ødelagt DNA som gjenspeiler våre felles aner med aper.
Tenk på argumentene til en BioEssays-artikkel, "Processed pseudogenes: A substrate for evolutionary innovation," nettopp publisert i september 2021 -lenke:
Implikasjonen av pseudogener i biologiske prosesser, inkludert nevrogenese, inflammatoriske responser og kreft, nødvendiggjør revisitasjon av forestillingen om at pseudogener er evolusjonært 'søppel'. Imidlertid er omfanget av pseudogen aktivitet fortsatt dårlig undersøkt, delvis kanskje på grunn av skjevheten som er iboende til begrepet "pseudogen", som forutsetter ikke-funksjonalitet. Videre har tekniske mangler hindret entydig forskjell på pseudogen aktivitet fra deres nesten identiske foreldremotparter.5 [Uthevelse tilføyd.]
Avisen forklarer at selv om pseudogener tradisjonelt har blitt "merket funksjonsløse i massevis", er sannheten at "forholdsvis lite er kjent" om dem:
Etter Ohnos antakelse om at de fleste dupliserte gener er bestemt for degenerasjon, konkluderte Jacq og medarbeidere at de identifiserte 5S ribosomale RNA 'pseudo'-genene var artefakter av evolusjon. Disse bemerkningene ga grunnlaget for et rammeverk som kategoriserer tilsynelatende defekte sekvenser med likhet med et annet gen som pseudogener. Genomikk-revolusjonen så deretter regioner av genomet med pseudogene kjennetegn merket funksjonsløs i massevis. Selv om pseudogener er nesten like mange som proteinkodende gener (henholdsvis 14 767, hvorav 72 % er prosessert, 24 % duplisert, 1,6 % duplisert, 2,4 % annet og 19 957), er det relativt lite kjent om bidraget til pseudogener til utviklingen av det menneskelige genom.
Bilde 7. RNA kodes ut fra DNA som mal
 Kristne intellektuelle som tror at pseudogener er ikke-funksjonelle søppel, adopterer antakelser direkte vokst ut av et evolusjonært paradigme - og dette er antagelser om et aspekt av genomet som vi faktisk vet veldig lite om. Verken kristne eller noen andre trenger å frykte at de fornekter god vitenskap ved å forutse funksjon for pseudogener, eller i det minste ved å være forsiktige med å hoppe på flertallsoppfatningen at de er genetisk 'søppel'.
Kristne intellektuelle som tror at pseudogener er ikke-funksjonelle søppel, adopterer antakelser direkte vokst ut av et evolusjonært paradigme - og dette er antagelser om et aspekt av genomet som vi faktisk vet veldig lite om. Verken kristne eller noen andre trenger å frykte at de fornekter god vitenskap ved å forutse funksjon for pseudogener, eller i det minste ved å være forsiktige med å hoppe på flertallsoppfatningen at de er genetisk 'søppel'.
Mange andre autoritative artikler argumenterer tilsvarende. En artikkel fra 2018 publisert i et Springer biokjemitidsskrift kommenterer:
I lang tid har pseudogener blitt betraktet som "søppel-DNA" som uunngåelig oppstår som et resultat av pågående evolusjonsprosess. Imidlertid indikerer eksperimentelle data innhentet i løpet av de siste årene at denne forståelsen av pseudogenenes natur ikke er helt korrekt, og mange pseudogener utfører viktige genetiske funksjoner. … pseudogener er integrerte komponenter i omfattende regulatoriske nettverk av samvirkende gener.7
Artikkelen fortsetter:
Utvikling av ny generasjons DNA-sekvensering (NGS) har ført til en uforutsett oppdagelse av det gjennomgripende transkripsjons-fenomenet. Selv om proteinkodende ekson-sekvenser bare representerer en liten brøkdel av dyre- og plantegenomer (<2% av det menneskelige genomet), blir flertallet av dem transkribert. Derfor er det ikke overraskende at pseudogen RNA (psRNA) er betydelig representert i de studerte transkriptomene. Dermed ser det ut til at omtrent 10 % av alle identifiserte (~15 000-18 000) menneskelige pseudogener er transkribert. I henhold til tilgjengelige eksperimentelle data kan transkriberte pseudogener deles inn i tre grupper: (1) universelt transkribert, (2) ikke-spesifikt transkribert og (3) transkribert på en strengt vevsspesifikk måte. Universelt uttrykte pseudogener er relatert til husholdningsgener som vanligvis har det største antallet pseudogener. Den andre gruppen av pseudogener, spesielt CYP4Z2P og derivater av Oct-4-, Connexin-43- og BRAF-gener, er sterkt uttrykt i en eller to vevstyper, men viser lave ekspresjonsnivåer i andre vev. Til slutt ble strengt vevsspesifikk transkripsjon vist for ~150 humane pseudogener, inkludert derivater av AURKA (nyrer) og RHOB (tarm) gener. Den vevsspesifikke karakteren til transkripsjon antyder at disse pseudogenene utfører vevsspesifikke funksjoner. Generelt har flertallet av transkriberte pseudogener blitt påvist i testiklene, mens muskelvevet har minst antall transkriberte pseudogener.
Det er viktig å sette pris på det du nettopp har lest: Vi ser allerede bevis på funksjon for omtrent 10 prosent av pseudogenene - noe som evolusjonister en gang trodde var umulig. Men mange pseudogener uttrykkes bare i spesifikke vev og til bestemte tider. Dette antyder at det kan være svært vanskelig å oppdage funksjonen og formålet til disse pseudogenene. Hvorfor? Fordi de kan være aktive bare sjelden i løpet av menneskets livssyklus. Du kan ikke holde et menneske i et laboratorium for en levetid for å se hva som skjer i hver vevstype. Som artikkelen forklarer:
Selv om det allerede har gått 40 år siden oppdagelsen av pseudogener, er undersøkelsen av disse genomiske komponentene med moderne metoder bare i begynnelsen. [Uthevelse lagt til.]
Likevel, som en artikkel i Annual Review of Genetics observerte: "pseudogener som har blitt hensiktsmessig undersøkt, viser ofte funksjonelle roller." (7) Likeledes uttaler en artikkel fra 2012 i RNA Biology at "pseudogener lenge ble betraktet som søppelgenomisk DNA", men "pseudogen regulering er utbredt i eukaryoter." Artikkelen konkluderer med at "studiet av funksjonelle pseudogener bare er i begynnelsen" og spår "flere og flere funksjonelle pseudogener vil bli oppdaget etter hvert som nye biologiske teknologier utvikles i fremtiden."
Disse nye teknologiene bærer allerede frukt. Litteraturen er full av artikler som rapporterer spesifikke funksjoner i deler av DNA, tidligere merket som 'pseudogenes' (9) ENCODE-prosjektet rapporterte over 850 menneskelige pseudogener som er "transkribert og assosiert med aktivt kromatin." (10) En artikkel i Molecular Biology and Evolution fant at "656 menneske-makak ortologe pseudogener ... blir transkribert." (11) Og en studie av det menneskelige proteomet publisert i Nature rapporterte "mer enn 200 peptider som er kodet av 140 pseudogener." (12)
Bilde 8. Anti-sense RNA kan hindre at mRNA oversettes til protein
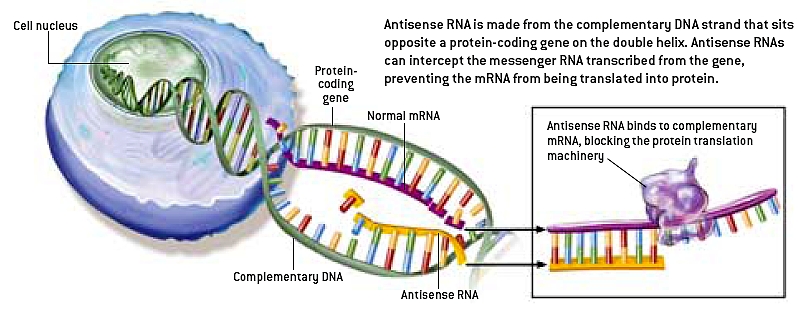 Massefunksjonalitet for pseudogener er ikke lenger vanskelig å tro. De kan gi funksjonelle proteiner, funksjonelle RNA-transkripter eller utføre en funksjon uten å produsere noen transkripsjon. Ved RNA-interferens gir et pseudogen et "anti-sense" RNA-transkript som ikke kan produsere et protein, men som kan binde seg med transkripsjoner av proteinkodende versjoner av genet. Når slik binding oppstår, kan ikke det proteinkodende transkripsjonen oversettes, noe som reduserer proteinproduksjonen.(13) Ved målmimik binder små RNA-er seg til et proteinkodende mRNA-transkript, og hemmer translasjon. Hvis et pseudogen produserer lokke-mRNA-transkripter som etterligner "mål"-sekvensen til de proteinkodende motstykkene, kan disse små RNA-ene binde seg til de pseudogene transkriptene i stedet. Dette forhindrer inhibering av translasjon, og øker proteinproduksjonen.(14) En av artiklene som er sitert ovenfor, lister opp flere typer funksjoner for pseudogen RNA, som de kaller "psRNA":
Massefunksjonalitet for pseudogener er ikke lenger vanskelig å tro. De kan gi funksjonelle proteiner, funksjonelle RNA-transkripter eller utføre en funksjon uten å produsere noen transkripsjon. Ved RNA-interferens gir et pseudogen et "anti-sense" RNA-transkript som ikke kan produsere et protein, men som kan binde seg med transkripsjoner av proteinkodende versjoner av genet. Når slik binding oppstår, kan ikke det proteinkodende transkripsjonen oversettes, noe som reduserer proteinproduksjonen.(13) Ved målmimik binder små RNA-er seg til et proteinkodende mRNA-transkript, og hemmer translasjon. Hvis et pseudogen produserer lokke-mRNA-transkripter som etterligner "mål"-sekvensen til de proteinkodende motstykkene, kan disse små RNA-ene binde seg til de pseudogene transkriptene i stedet. Dette forhindrer inhibering av translasjon, og øker proteinproduksjonen.(14) En av artiklene som er sitert ovenfor, lister opp flere typer funksjoner for pseudogen RNA, som de kaller "psRNA":
-psRNA deltar i reguleringen av foreldrenes genuttrykk på både transkripsjons- og translasjonsnivå. Sensestrengen av DNA fungerer som en mal for syntesen av sense-mRNA (sRNA), mens transkripsjon av den antiparallelle strengen produserer komplementær antisense-RNA (asRNA). Siden både sRNA og asRNA syntetiseres under pseudogen transkripsjon, vil vi videre diskutere den funksjonelle relevansen til pseudogen-transkripsjonene under hensyntagen til denne strukturelle forskjellen.
-Funksjoner til sRNA. Typisk skiller sekvenser av pseudogene sRNA-er seg bare litt fra foreldregen-mRNA. På grunn av den høye likheten deler de miRNA-bindingssteder (miRNA-responselementer, MRE), hvis binding til miRNA sikrer regulatorisk funksjon av disse RNA-molekylene i både kjernen og cytoplasmaet. … Jo høyere pseudogen transkripsjonsaktivitet, desto høyere er antallet miRNA-molekyler som binder seg til dets sRNA, noe som tømmer deres intracellulære pool og reduserer undertrykkelse av det parentale genuttrykket, dvs. forholdet mellom mengdene av gen- og pseudogentranskripsjoner justerer genuttrykket ….
-Funksjoner til asRNA. Antisense-transkripsjon er funnet i alle studerte levende organismer. NGS-dataene (neste generasjons sekvensering) tyder på at >70 % RNA-er i muse-transkriptomet overlapper med komplementære sekvenser, og representerer dermed naturlige antisense-transkripter. Det samme gjelder for det menneskelige transkriptomet. Hos mus og mennesker er flertallet av asRNA-er strengt bevart, noe som indikerer deres funksjonelle relevans, noe som er eksperimentelt bekreftet. Nå er mange funksjoner til asRNA-er identifisert. asRNA kan påvirke målgenene deres både i cis og i trans. Noen av disse aktivitetene er også observert for pseudogene asRNAer.
Bilde 9.Kodoner og antikodoner (aminosyrer)
 -Pseudogene asRNA-er i dannelsen av duplekser med foreldregen-sRNA. Oppdagelsen av involveringen av pseudogene asRNA-er i inhiberingen av translasjon av foreldregen-mRNA-er var en av de første indikasjonene på deres funksjonelle rolle i levende organismer. Denne mekanismen realiseres gjennom dannelsen av RNA-RNA-duplekser mellom svært homologe regioner av pseudogenet asRNA og translatert mRNA med påfølgende undertrykkelse av de tilsvarende gener på translasjonsnivå ....
-Pseudogene asRNA-er i dannelsen av duplekser med foreldregen-sRNA. Oppdagelsen av involveringen av pseudogene asRNA-er i inhiberingen av translasjon av foreldregen-mRNA-er var en av de første indikasjonene på deres funksjonelle rolle i levende organismer. Denne mekanismen realiseres gjennom dannelsen av RNA-RNA-duplekser mellom svært homologe regioner av pseudogenet asRNA og translatert mRNA med påfølgende undertrykkelse av de tilsvarende gener på translasjonsnivå ....
-Pseudogene asRNA-er i dannelsen av korte interfererende RNA-er (siRNA). Pseudogene asRNA-er som deltar i dannelsen av RNARNA-duplekser kan gi opphav til siRNA-er. Strukturen og virkningsmekanismen til regulatoriske siRNA-er ligner på miRNA-er...
-Pseudogene asRNA-er i generering av piRNA-er. Pseudogen-kodede RNA-er som interagerer med PIWI-proteiner (piRNA-er) ble nylig funnet blant korte ikke-kodende RNA-er i sædceller fra mennesker og dyr og kimlinjeceller ...
-Pseudogen asRNA i reguleringen av transkripsjon av målgener. Nå er rollen til lncRNA i epigenetiske genommodifikasjoner akkompagnert med etablering av genekspresjonsmønstre i celler blitt demonstrert og omfattende studert. Lignende egenskaper ble også observert for noen pseudogene RNA...(15)-
Mange andre artikler har advart mot å avfeie pseudogener som rent søppel. En artikkel fra 2012 i 'Science Signaling' bemerket at selv om "pseudogener lenge har blitt avvist som søppel-DNA", har nyere fremskritt fastslått at "DNA til et pseudogen, RNA transkribert fra et pseudogen, eller proteinet oversatt fra et pseudogen kan ha flere, forskjellige funksjoner og at disse funksjonene kan påvirke ikke bare deres foreldregener, men også ikke-relaterte gener.-" (16) Deres homologi til de p roteinkodende versjonene av genet er nødvendig for denne regulatoriske funksjonen, som "pseudogener, på grunn av deres høysekvens-homologi, kan fungere som legitime bona fide mikroRNA-konkurrenter, og dermed aktivt konkurrere med deres forfedres proteinkodende gener om den samme poolen av mikroRNA gjennom sett med konserverte MRE-er [mikroRNA-responselementer]." (17) 'Science Signaling' artikkelen konkluderer med at "pseudogener har dukket opp som en tidligere ukjent klasse av sofistikerte modulatorer av genuttrykk." En artikkel fra 2011 i tidsskriftet RNA er enig:
roteinkodende versjonene av genet er nødvendig for denne regulatoriske funksjonen, som "pseudogener, på grunn av deres høysekvens-homologi, kan fungere som legitime bona fide mikroRNA-konkurrenter, og dermed aktivt konkurrere med deres forfedres proteinkodende gener om den samme poolen av mikroRNA gjennom sett med konserverte MRE-er [mikroRNA-responselementer]." (17) 'Science Signaling' artikkelen konkluderer med at "pseudogener har dukket opp som en tidligere ukjent klasse av sofistikerte modulatorer av genuttrykk." En artikkel fra 2011 i tidsskriftet RNA er enig:
Bilde 10. mikro-RNA kan hindre oversettelse til protein
-Pseudogener har lenge vært merket som "søppel" DNA, mislykkede kopier av gener som oppstår under utviklingen av genomer. Nyere resultater utfordrer imidlertid denne betegnelsen; faktisk ser det ut til at noen pseudogener har potensialet til å regulere deres proteinkodende fettere. (18)
Som en artikkel fra 2013 i eLife sier: "Pseudogener antas å være inaktive gensekvenser, men nyere bevis på omfattende pseudogen transkripsjon reiste spørsmålet om potensiell funksjon." (19) En annen artikkel i eLife fra 2015 sier: "Bruk av en ny bio-informatisk metode for å analysere ribosom-profileringsdata viser vi at 40 % av lncRNA-er og pseudogene RNA-er uttrykt i humane celler blir oversatt [til proteiner]."(20) Artikkelen fortsetter:
-Eksperimentene viser at tusenvis av ikke-kodende RNA-er i det menneskelige genomet faktisk er oversatt. Dette er mange flere enn forventet og representerer omtrent 40 % av lncRNA-ene og pseudogene RNA-ene, og 35 % av uoversatte regioner i messenger-RNA.
Sekvensbevaring kan også innebære pseudogen funksjonalitet selv når en spesifikk funksjon ennå ikke er oppdaget. En studie fra 2014 i Proceedings of the National Academy of Sciences sammenlignet pseudogener hos mennesker, C. elegans og fruktfluer. Den fant direkte bevis på funksjon så vel som konservert sekvensering, noe som tyder på funksjon: "Samlet sett identifiserer vi et bredt spekter av biokjemisk aktivitet for pseudogener, med flertallet i hver organisme som viser varierende grad av delvis aktivitet. Spesielt identifiserer vi en konsistent mengde transkripsjon (~15%) på tvers av alle arter, noe som antyder en enhetlig nedbrytningsprosess. Vi ser også et jevnt forfall av pseudogen promoter-aktivitet i forhold til deres kodende motparter og identifiserer en rekke pseudogener med konserverte oppstrømssekvenser og aktivitet, noe som antyder potensielle regulatoriske roller.-" (21) En artikkel fra 2011 i Cell antyder på samme måte funksjoner i mange pseudogener: Sekvenserings-innsats har avslørt ~19 000 pseudogener hos mennesker, hvorav mange er transkribert og ofte godt bevart, noe som tyder på at det eksisterer selektivt press for å opprettholde pseudogener.-"(22)
En av de mest dyptgripende og fremtredende uttalelsene om emnet kom i en artikkel fra 2020 i Nature Reviews Genetics. Denne artikkelen advarte om at pseudogen funksjon er "for tidlig avvist" på grunn av "dogmer", der "Den dominerende begrensningen i å fremme etterforskningen av pseudogener nå ligger i fangstene til den rådende tankegangen at pseudogene regioner, i seg selv, er ikke-funksjonelle." I stedet finner artikkelen at "Der pseudogener har blitt studert direkte, er de ofte funnet å ha kvantifiserbare biologiske roller." Noen av disse funksjonene er "proteinbaserte", noe som betyr at pseudogenet genererer et funksjonelt protein. Men andre funksjoner kan være "RNA-baserte" eller "DNA-baserte." For eksempel antar evolusjonister vanligvis at et pseudogen som ikke produserer et protein, ikke kan være funksjonelt. Men papiret observerer at pseudogener som ikke kan oversettes til et protein fortsatt kan ha en funksjon, gjennom RNA-transkripsjonen deres:
-Mange pseudogener inneholder en frekvens av mutasjoner som gjør dem usannsynlig å bli (eller ute av stand til å bli) oversatt til proteiner. Imidlertid utelukker ikke slike mutasjoner nødvendigvis pseudogener fra å utføre en biologisk funksjon.
En rekke andre ikke-transkripsjonelle funksjoner er dokumentert i artikkelen, inkludert stabiliserende kromosomer, mediering av transkripsjonsspleising og regulering av rekombinasjon. I mange tilfeller ser derfor kopiantall av pseudogener ut til å ha funksjonell betydning, der avvik fra den normale genetiske tilstanden forårsaker sykdom. De spår: "Det forventes at ytterligere koblinger mellom menneskelige pseudogene polymorfismer og komplekse sykdommer vil bli identifisert i de kommende årene."
Det typiske svaret fra evolusjonister vil være at disse eksemplene bare er isolerte sjeldne tilfeller, og at hoveddelen av pseudogener likevel er useriøse. Forfatterne av Nature Reviews Genetics-artikkelen - som ikke gir noen indikasjon på sympati for intelligent design - er klar over denne innvendingen. De sier følgende som direkte avvisning til det:
Bilde 11. Retrogen -fungerende pseudogen
 -Eksemplene på pseudogen funksjon som er utdypet her, bør ikke innebære at pseudogen funksjonalitet
-Eksemplene på pseudogen funksjon som er utdypet her, bør ikke innebære at pseudogen funksjonalitet
annsynligvis vil være begrenset til isolerte tilfeller. Minst 15 % av pseudogenene er transkripsjonelt aktive på tvers av tre phyla, hvorav mange er proksimale til bevarte regulatoriske regioner. Det er anslått at minst 63 nye humanspesifikke proteinkodende gener ble dannet ved retro-transposisjon siden avviket fra andre primater. Tallrike 'retrogener' fortsetter å bli anerkjent som funksjonelle proteinkodende gener, i stedet for pseudogener på tvers av arter. Massespektrometri med høy gjennomstrømning og ribosomal profilering har identifisert hundrevis av pseudogener som er oversatt til peptider. Selv om funksjonene til disse peptidene gjenstår å bli eksperimentelt bestemt, illustrerer slike eksempler utfordringen med å underbygge en gen-pseudogen dikotomi.
De fortsetter: "Ettersom overfloden av slike [ikke-kodende-DNA]-ervervede funksjoner ikke ser ut til å være et spesielt sjeldent eller isolert fenomen, vil det virke upassende å ta standardperspektivet at behandlede pseudogener er funksjonsløse. I stedet er det sannsynlig at pseudogenholdige områder av genomet har viktige biologiske funksjoner som ennå ikke er avslørt."
Bilde 12. CRISPR gen-editering hos mennesket
 For tiden bemerker imidlertid forfatterne, "delvis på grunn av den eksperimentelle utfordringen med å undersøke deres funksjon og uttrykk, er pseudogener vanligvis ekskludert fra genom-omfattende funksjonelle screenings- og uttrykks-analyser." Med andre ord, en av hovedårsakene til at vi ikke finner funksjon for pseudogener, er fordi vi ikke leter etter det. En komplikasjon er at pseudogen transkripsjon viser "celletype spesifisitet og dynamisk uttrykk" - noe som betyr at de bare kan transkriberes på bestemte steder til bestemte tider. Dette er desto større grunn til ikke å anta at mangel på bevis for funksjonen til et pseudogen er bevis på at pseudogenet ikke har noen funksjon! Det kan meget sannsynlig være funksjonelt i en celletype eller en situasjon som vi bare ikke har undersøkt ordentlig ennå. Som de sa det, "Bruk av analyser som er dårlig egnet til analyse av pseudogener, har uten tvil hindret klargjøring av deres biologiske roller." Men de er håpefulle: "CRISPR-baserte tilnærminger, nøye anvendt, har potensial til å revolusjonere vår evne til å dissekere funksjonene til pseudogener."
For tiden bemerker imidlertid forfatterne, "delvis på grunn av den eksperimentelle utfordringen med å undersøke deres funksjon og uttrykk, er pseudogener vanligvis ekskludert fra genom-omfattende funksjonelle screenings- og uttrykks-analyser." Med andre ord, en av hovedårsakene til at vi ikke finner funksjon for pseudogener, er fordi vi ikke leter etter det. En komplikasjon er at pseudogen transkripsjon viser "celletype spesifisitet og dynamisk uttrykk" - noe som betyr at de bare kan transkriberes på bestemte steder til bestemte tider. Dette er desto større grunn til ikke å anta at mangel på bevis for funksjonen til et pseudogen er bevis på at pseudogenet ikke har noen funksjon! Det kan meget sannsynlig være funksjonelt i en celletype eller en situasjon som vi bare ikke har undersøkt ordentlig ennå. Som de sa det, "Bruk av analyser som er dårlig egnet til analyse av pseudogener, har uten tvil hindret klargjøring av deres biologiske roller." Men de er håpefulle: "CRISPR-baserte tilnærminger, nøye anvendt, har potensial til å revolusjonere vår evne til å dissekere funksjonene til pseudogener."
Artikkelen advarer om at det er mange tilfeller der DNA, som ble avvist som pseudogent søppel, senere ble funnet å være funksjonelt: "med et økende antall tilfeller av pseudogen-annoterte regioner som senere ble funnet å vise biologisk funksjon, er det en voksende risiko for at disse regionene av genomet blir for tidlig avvist som pseudogene og derfor ansett som funksjonsløse.-" (23)
Pseudogen feil fra fortiden
Det er presedens for at kristne, som for tidlig avviser pseudogener som ødelagt DNA, for senere å bli bevist feilaktige. Det samme skjedde i tidligere, høyprofilerte situasjoner. Under Kitzmiller mot Dover-rettssaken i 2005, vitnet den ledende teistiske evolusjonsbiologen Kenneth Miller om at det menneskelige beta-globin-pseudogenet er 'ødelagt' fordi "det har en rekke molekylære feil som gjør genet ikke-funksjonelt-". Siden mennesker, sjimpanser og gorillaer deler "matchingsfeil" i pseudogenet, sa han til retten, "leder dette oss til bare én konklusjon … at disse tre artene deler en felles stamfar." (24) Han tok feil. En studie fra 2013 i Genome Biology and Evolution rapporterte at beta-globin-pseudogenet er funksjonelt. (25)
Mennesker har seks kopier av beta-globingenet. Fem produserer beta-globinproteiner, men den sjette, den pseudogene kopien, har et for tidlig stoppkodon som hindrer riktig oversettelse. Forskerne sammenlignet alle seks gener på tvers av mennesker og sjimpanser, og fant ut at beta-globin-pseudogenet viser færre forskjeller enn det som ville vært forventet, hvis det var ikke-funksjonelle og akkumulerte tilfeldige mutasjoner med en konstant hastighet. Denne "konserverte" sekvensen antyder at beta-globin-pseudogenet har en selekterbar funksjon, noe som gjør det mindre tolerant overfor mutasjoner. Beta-globin-pseudogenets manglende evne til å produsere et translaterbart RNA-transkript utelukker ikke at det er funksjonelt. Forskerne argumenterer for at pseudogenet fungerer som en av/på-bryter, som regulerer uttrykket av proteinkodende beta-globin-gener under embryonal utvikling.
Lære leksjonen om Pseudogener på nytt
Til tross for alle disse bevisene, antar de fleste evolusjonister fortsatt at pseudogener er søppel-DNA. Dette forblir 'konsensu'-synet. I mellomtiden ser mange kristne intellektuelle ikke antakelsene eller setter pris på det raskt voksende beviset på pseudogen funksjon. De ser bare konsensusen, og mener vi ikke må motsette oss den. Men trendlinjen til bevisene viser at pseudogener ikke bør antas å være 'ødelagt' DNA. For å oppsummere er det mange grunner til å ta i bruk en "vent og se" tilnærming til pseudogener:
*Pseudogen ikke-funksjonalitet er en antagelse som stammer fra en evolusjonær tankegang, ikke en konklusjon fra nøye studier av bevisene.
*Faktisk har pseudogener knapt blitt studert - hovedsakelig, fordi teknologi ennå ikke er utviklet for å studere dem. Vår mangel på teknisk evne til å studere pseudogener stammer direkte fra utbredte evolusjonære antakelser om at de ikke gjør noe, og at vi ikke bør kaste bort tiden vår på å prøve å finne ut hva de gjør.
*Selv om det var helt uventet under en evolusjonær 'søppel' DNA-tankegang, har mange spesifikke eksempler på pseudogen funksjon blitt funnet. I andre tilfeller har pseudogen funksjon blitt utledet basert på biokjemisk aktivitet av pseudogent DNA eller bevaring av sekvens.
*Dette beviset på pseudogen funksjon er ikke overraskende, fordi vi vet at det er mange måter pseudogener kan ha funksjon på: Pseudogener kan gi funksjonelle proteiner, funksjonelle RNA-transkripsjoner eller utføre en funksjon uten å produsere noen transkripsjon. Mange pseudogene RNA-er ser ut til å ha genregulerende funksjoner, ofte i en epigenetisk kontekst.
 *Vi er bare i begynnelsen av å utvikle teknologi for å forstå pseudogener. Mange pseudogener kan være aktive bare i én vevstype eller i visse stadier av livssyklusen, noe som gjør deres funksjoner vanskelig å oppdage. Så det er ikke overraskende at vi har få eksempler på funksjonelle pseudogener. Men etter hvert som vi utvikler bedre teknologier for å studere pseudogener, viser funksjonalitet seg å være mer og mer utbredt.
*Vi er bare i begynnelsen av å utvikle teknologi for å forstå pseudogener. Mange pseudogener kan være aktive bare i én vevstype eller i visse stadier av livssyklusen, noe som gjør deres funksjoner vanskelig å oppdage. Så det er ikke overraskende at vi har få eksempler på funksjonelle pseudogener. Men etter hvert som vi utvikler bedre teknologier for å studere pseudogener, viser funksjonalitet seg å være mer og mer utbredt.
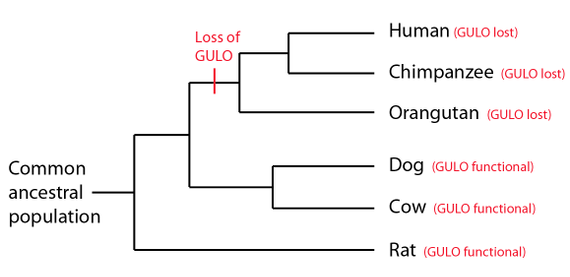
Bilde 13. Tap av GULO-gen -devoluert?
Riktignok forblir de nøyaktige funksjonene til mange pseudogener ukjente. For eksempel er vitamin C "gulo" pseudogen, et enhetlig pseudogen som deles av mennesker og mange primater, ennå ikke kjent for å ha funksjon. Det er derfor det har blitt et populært argument mot intelligent design blant tilhengere av TE/EC. Imidlertid har selv dette pseudogenet blitt foreslått å være aktivt in utero. Før 2013 kunne det samme ha blitt sagt om beta-globin-pseudogenet, som nå forstås å ha funksjon.
Kristne intellektuelle som antar at pseudogener er ødelagt DNA, satser på en hest som knapt har begynt å løpe sitt løp - og når hesten begynner å løpe, ser vi at den ikke gjør det så bra. Akkurat som en artikkel i Nature Reviews Genetics sitert ovenfor hevdet at pseudogen funksjon er "prematurt avskjediget", avviser noen ortodokse kristne doktriner for tidlig. Selv for de som ikke føler seg klare til å støtte oppfatningen om at pseudogener er funksjonelle, virker i det minste en agnostisk "vent og se"-tilnærming fornuftig, gitt mangelen på kunnskap og trendlinjen til bevisene.
Bilde 14. Casey Luskin -opphavsmann til artikkelen
 La oss snakke om den trendlinjen. Pseudogener er bare én type DNA som antas å være "søppel". Tilbake på 1990-tallet og begynnelsen av 2000-tallet ble over 90 prosent av genomet vanligvis antatt å være genetisk søppel - et syn som nå avvises av de fleste biologer. Intelligent design-teoretikere spådde at mye av søppel-DNA ville vise seg å ha funksjon. De hadde rett: En revolusjon har i stor grad overtatt biologisk tenkning. Selv om det fortsatt er mye vi ikke vet om genomet, er trendlinjen sterkt mot 'søppelsynet'. Hvis du ikke tror meg, se på den oransje linjen i denne grafen fra tidsskriftet Nature. Den viser den raske, jevne eksponentielle hastigheten som funksjoner blir oppdaget for ikke-kodende genetiske elementer - som Nature sier tidligere ble kalt 'søppel-DNA' -lenke.
La oss snakke om den trendlinjen. Pseudogener er bare én type DNA som antas å være "søppel". Tilbake på 1990-tallet og begynnelsen av 2000-tallet ble over 90 prosent av genomet vanligvis antatt å være genetisk søppel - et syn som nå avvises av de fleste biologer. Intelligent design-teoretikere spådde at mye av søppel-DNA ville vise seg å ha funksjon. De hadde rett: En revolusjon har i stor grad overtatt biologisk tenkning. Selv om det fortsatt er mye vi ikke vet om genomet, er trendlinjen sterkt mot 'søppelsynet'. Hvis du ikke tror meg, se på den oransje linjen i denne grafen fra tidsskriftet Nature. Den viser den raske, jevne eksponentielle hastigheten som funksjoner blir oppdaget for ikke-kodende genetiske elementer - som Nature sier tidligere ble kalt 'søppel-DNA' -lenke.
Ikke sats på feil hest. Vær forsiktig med å kaste bort tusenårs gamle doktriner fordi du blir fortalt at konsensus krever at du må tro at pseudogener er 'ødelagte' gener som er arvet fra våre apelignende forfedre. Konsensus kan si dette, men bevisene gjør det ikke. Hvis du ønsker å stille spørsmål ved eller fullstendig bryte med konsensus om pseudogen funksjonalitet, er bevisene der for å støtte deg. Ha mot til å følge bevisene hvor enn de fører.
Referanser -se nederst her.
Oversettelse og bilder ved Asbjørn E. Lund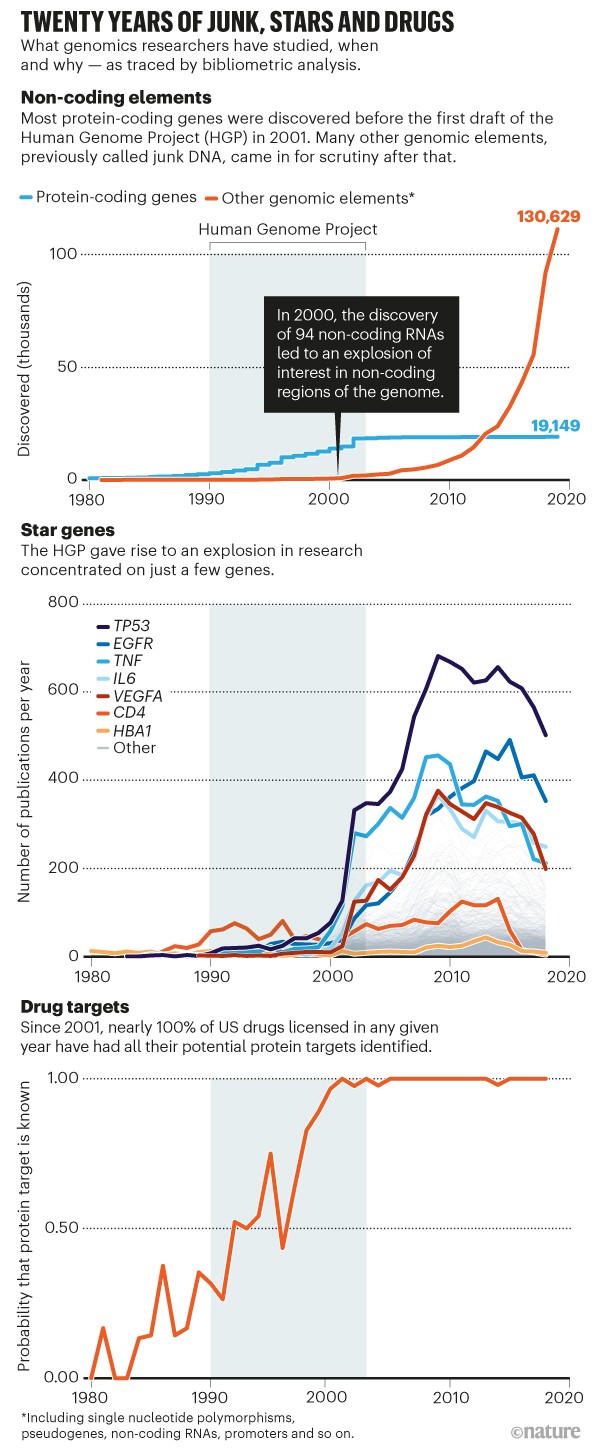{kind=link}